Тематический план
Общее
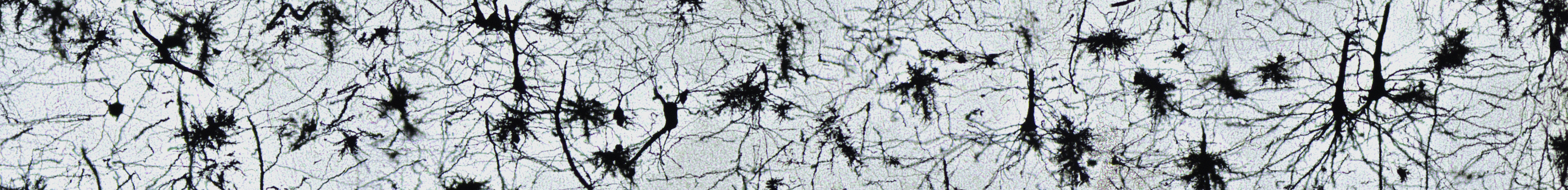
Внутренний пирамидальный слой моторной коры головного мозга. Препарат кафедры гистологии, цитологии и эмбриологии Казанского ГМУ.
Дистанционный курс обучения по модулю "Нервная система" предназначен для студентов 1 курса, обучающаяся по интегрированному учебному плану по специальности 31.05.01 «Лечебное дело».
Разработчики:
- профессор кафедры гистологии, цитологии и эмбриологии, д.м.н. Челышев Юрий Александрович.
- доцент кафедры гистологии, цитологии и эмбриологии, к.м.н. Измайлов Андрей Александрович
Мотивация:
Дисциплина «Гистология, эмбриология, цитология», как и другие фундаментальные дисциплины, формирует научное представление о закономерностях развития, строения и жизнедеятельности организма человека. Изучение "Гистологии, эмбриологии, цитологии", как одной из базисных дисциплин в медицинском образовании, является обязательной частью образовательного процесса для студентов младших курсов, способствует формированию целостного восприятия структуры и функционирования органов и систем человеческого организма. Изучение дисциплины «Гистология, эмбриология, цитология» создает базис морфологических знаний, необходимый для успешного освоения других медико-биологических, а также клинических дисциплин.
Неделя 3. Ганглии, сплетения, нервы
Неделя 12.
Неделя 13. Автономная (вегетативная) нераная система (АНС)
Неделя 14. Орган зрения
Неделя 18. Кожа
